The Razor's Edge in the Grand Arc of Life
Digital twins in HSML and AGI, attractor navigation, and the criticality of the life process as an Omega trajectory from conception to mortality.

Digital twins, attractors, and the criticality of life
Life is not a static state. It is a trajectory held near a narrow operating zone where energy flow can generate adaptive information without falling into silence or runaway disorder.
An external model that observes a system, predicts its future states, and guides action.
An internal phase-space representation that sustains the organism through continuous correction.
A nested attractor architecture maintaining criticality across genetic, neural, metabolic, and homeostatic scales.
The twin principle
The modern idea of a digital twin usually begins in engineering. A machine, robot, patient, factory, or city is mirrored in software. The twin observes the system, receives data from it, predicts future states, and guides action.
HSML extends this idea into the deeper grammar of life. In HSML, a twin is not merely a digital copy. It is a phase-space representation that transacts information with the physical body. It encodes where the system is, what it is coupled to, what it can become, and whether a transition into a new attractor regime is worth the energetic cost.
Biology discovered this principle before technology named it. The genome is the deepest twin: a compressed phase-space description of the organism's developmental attractor landscape. Gene regulatory networks execute that landscape. Cell fates are not assembled mechanically. They crystallise as attractor states in gene-expression space.
Attractor layers
Development is a sequence of B2 crossings. Each crossing is an emergence event. A stem cell moves through possibility and commits to a fate. An organ system forms. A neural architecture stabilises. A body becomes viable.
The nervous system repeats the same mathematics at a radically faster timescale. Neural attractors are GRN attractors running in milliseconds. A percept, a memory, a decision, and a conscious state are each temporary stabilisations of neural activity inside basins of attraction.
The cortex is a hierarchy of nested strange attractors: columns within areas, areas within networks, and networks within hemispheres. Each level carries its own Omega and crosses its own threshold.

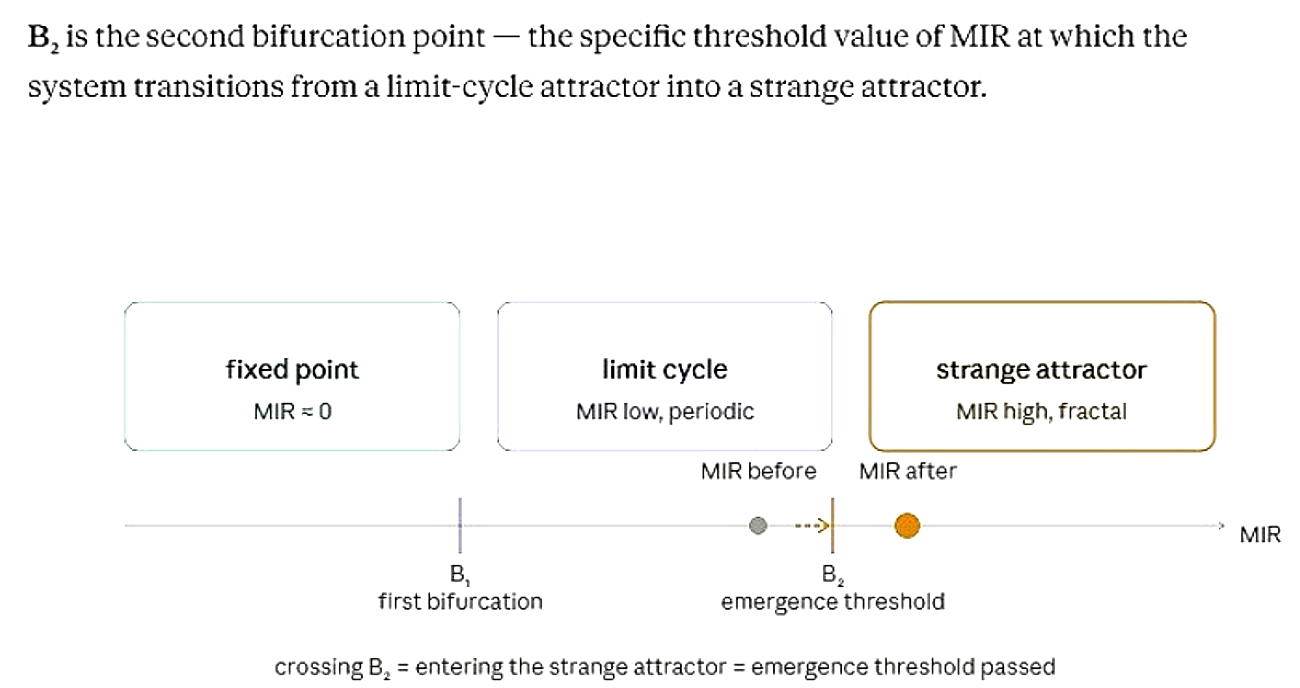
B2 as the emergence threshold
MIR is the Mutual Information Rate — how fast information is being exchanged between subsystems (say, kinetic and potential energy, or two brain regions). It is a continuous quantity that grows as the system becomes more chaotic and correlated.
B2 is the second bifurcation point — the specific threshold value of MIR at which the system transitions from a limit-cycle attractor into a strange attractor.
Below B2, the system may remain periodic and predictable, operating as a limit cycle. Once MIR rises past B2, the attractor changes type: the system enters a strange attractor, where correlations become long-range, fractal, and multi-scale.
So "MIR crosses B2" means the information exchange rate between the system's parts has become high enough to tip the system into the strange-attractor regime. That crossing is the moment emergence occurs: collective behaviour appears, long-range correlations form, and the system starts producing information across scales simultaneously.
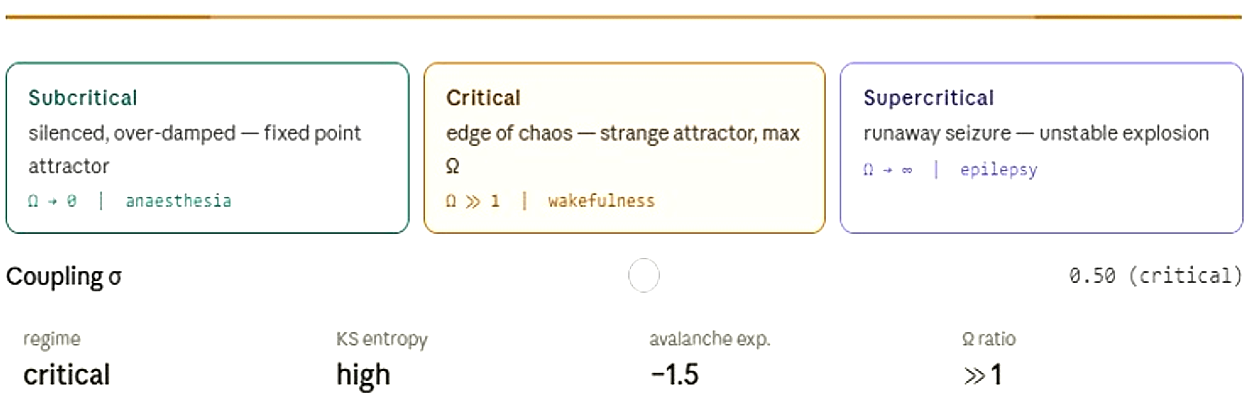
The Brain stays at the edge

Biological Master Reference Zero Point
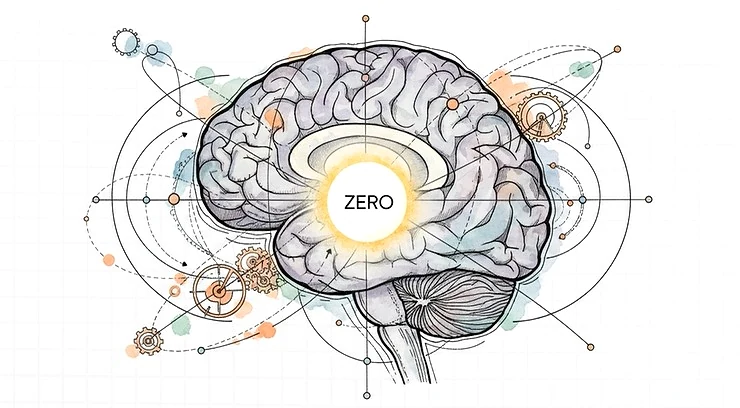
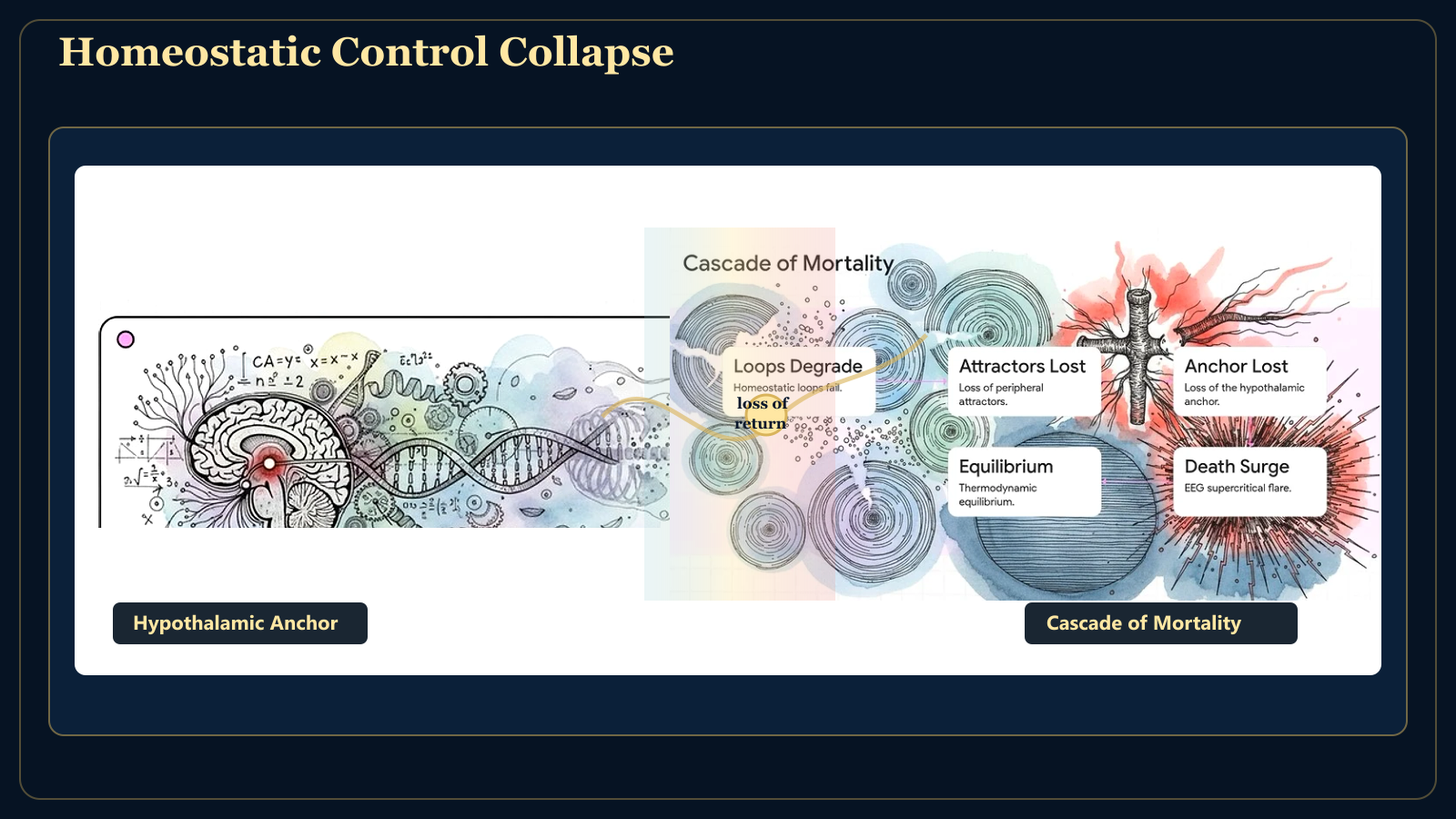
At the centre of this regulatory architecture sits the hypothalamus. It anchors hunger, thirst, temperature, glucose, osmolality, sleep, stress tone, endocrine rhythm, and autonomic balance.
It is not the richest information processor in the brain. It is the deepest reference point. In fact the Hypothalamus is the ground Zero and Initial Origin of intelligence in evolution of brain as ANS and BNS fuse , and this is the very first locus point of brain as we know it. Every higher excursion into cognition, emotion, imagination, social life, and symbolic intelligence remains ultimately tethered to the question the hypothalamus enforces: can this body continue?
Omega and mortality
Read across the lifespan, life becomes a single Omega curve. From conception to birth, GRN attractors crystallise and Omega rises. Through infancy and development, attractor layers multiply and coordinate. At peak vitality, the organism sustains criticality: maximum adaptive information per metabolic joule.
In ageing, homeostatic loops weaken, attractor basins flatten, and Omega begins to drift. In decline, return becomes uncertain. At death, the system can no longer maintain criticality across nested scales.
Mortality is therefore not simply silence. The final transition may first appear as supercritical information loss. As inhibitory and homeostatic control collapse, MIR may spike briefly. But this spike is not renewed life. It is the last uncontrolled cascade before the strange attractor dissolves.
Then MIR falls, Omega falls, KS entropy tends toward zero, and phase space contracts to thermodynamic equilibrium. Chaos was the path. Silence is the destination.