In our present generation folklore : Anesthesiologist can be compared to a highly skilled Ace Pilot ,who is navigating an emergent life process miracle known as Counciousness : taking off (Induction of Anesthesia ), keeping flight course till desired end target point is reached , well within homeostatic trajectory (maintainance of Anesthesia) ... and returning back to same body portal in life with comfortable Landing....(return of full conciousness ).
Or, Even maybe, more comparable to Submarine commander who dives down the vehicle known as body in the depth of ocean abyss ... then keeps peaceful, silent , but safe , ..ends dive as planned ... and emerge back to the normal mayhem of Councious life.
The Ace Pilot & The Submarine Commander
But ... MY take is .. Magical ....
: YOUR CHOICE to pick !!! .
Preamble
The Anesthesiologist's Framework
Anesthesia — General (GA), Relevant Against "Neural Attractors"
Neural Attractors are used reversibly — within safe limits of life control processes.
A) Targets for the Anesthesiologist
Sensory Data → Arousal (Wake) → Awareness (Understand) → Motor Response
The four targets: Awareness, Pain, Memory & Reflexes, Homeostasis (Metabolism).
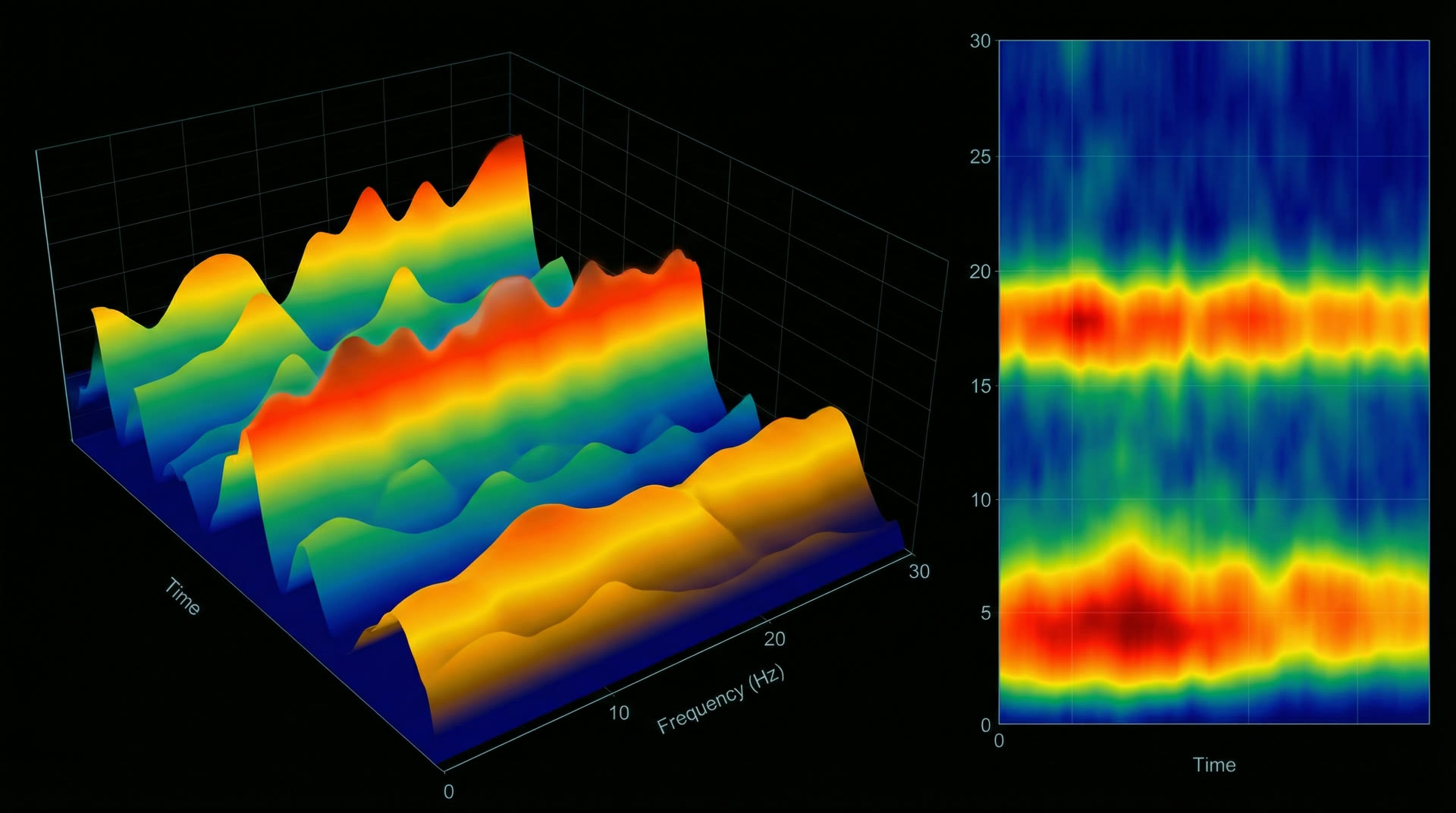
Waking human EEG comprises nested oscillatory bands: delta (0.5–4 Hz, 20–200 µV in deep sleep), theta (4–8 Hz, prominent in hippocampus at 15–25 µV scalp), alpha (8–13 Hz, 30–50 µV posterior), beta (13–30 Hz, 5–30 µV), and gamma (30–100 Hz, 2–10 µV). These are not independent — they are organised through cross-frequency coupling (CFC).
The primary CFC mechanism is phase-amplitude coupling (PAC): the phase of a slow oscillation modulates the amplitude of a fast oscillation. Canolty et al. (2006, Science, 313:1626–1628) demonstrated robust theta-gamma PAC in human electrocorticography (ECoG): gamma amplitude peaks at specific theta phases, with modulation index (MI) values of 0.01–0.03. Each theta cycle (125–250 ms) carries 5–8 gamma packets (~25 ms each), creating a temporal multiplexing scheme where each gamma burst encodes a distinct neural representation.
Thalamocortical loop circuits (most important & relevant circuits for consciousness): Thalamocortical relay neurons in the ventral posterior, lateral geniculate, and mediodorsal nuclei generate alpha-frequency oscillations via an intrinsic mechanism: the interplay between T-type calcium channels (Ca_v3.1) and hyperpolarization-activated cation channels (HCN/I_h) produces 8–13 Hz rhythmic bursting when the membrane potential is in the range −60 to −65 mV (Llinás & Steriade 2006, J. Neurophysiol., 95:3297–3308). This thalamic alpha gates cortical processing by cyclically inhibiting and disinhibiting cortical neurons, creating 100-ms windows of enhanced excitability (Haegens et al. 2011, PNAS, 108:4561–4566).
Disruption at any level of this hierarchy — loss of gamma coherence, decoupling from theta, disruption of thalamic alpha — alters or abolishes consciousness. General anesthesia provides the most controlled disruption, allowing systematic investigation of which oscillatory features are necessary for consciousness.
Nested oscillatory hierarchy — delta through gammaCompressed Spectral Array (CSA) — brain function monitor display during anesthesia
II
What Anesthesia Does
Not Silence but Disconnection · Metabolic Data · Feedforward vs Feedback
Under general anesthesia, cerebral metabolic rate of oxygen (CMRO₂) decreases by only 20–40% from the waking value of ~3.5 mL O₂/100g/min (Kaisti et al. 2003, Anesthesiology, 99:603–613). Cerebral blood flow (CBF) drops roughly proportionally. Neurons continue to fire — single-unit recordings in anesthetised animals show sustained spontaneous activity at 0.5–5 Hz in cortical neurons (Steriade et al. 2001, J. Neurophysiol., 85:1969–1985). The brain is not "turned off."
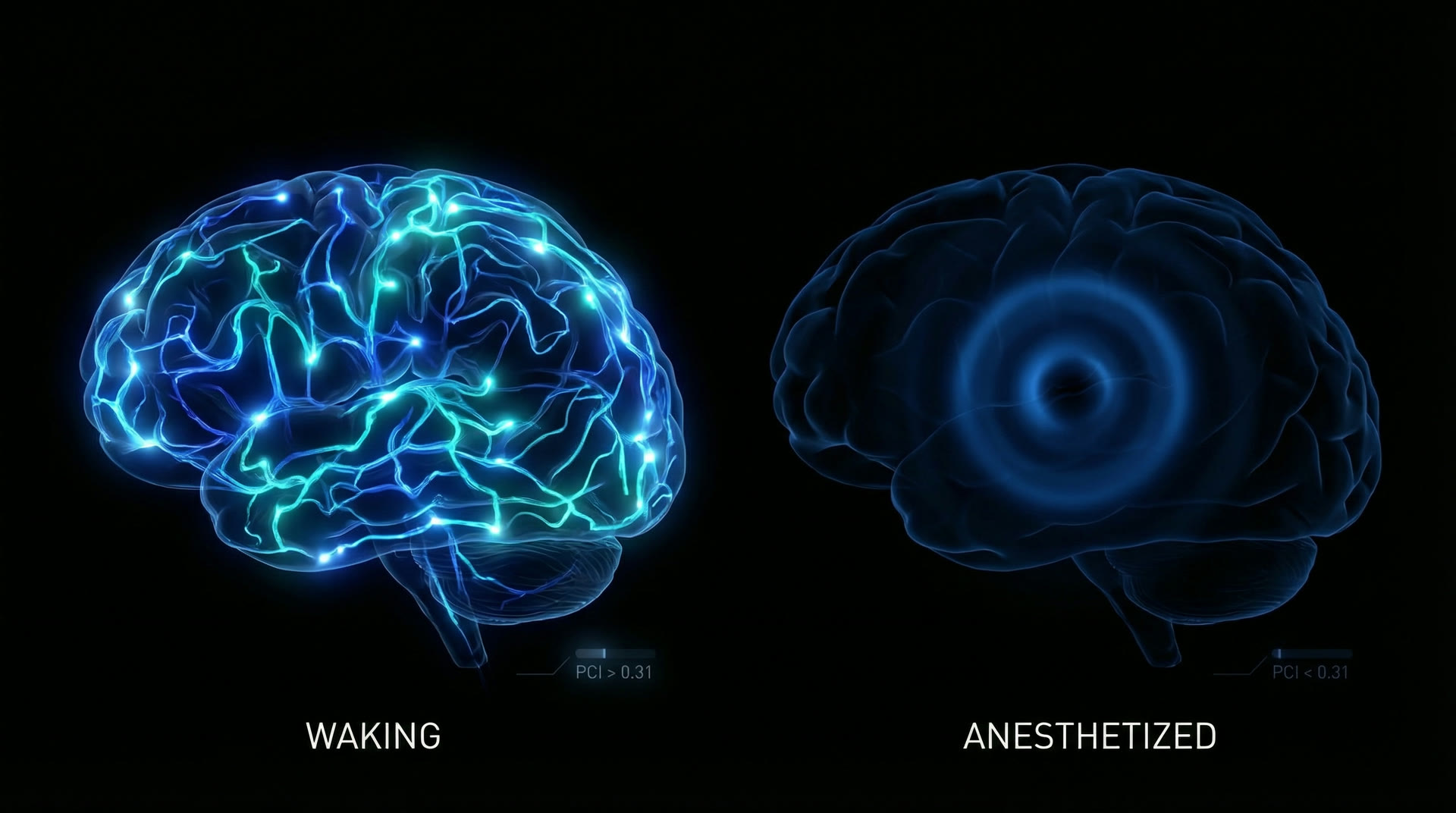
What changes is inter-regional communication. Ferrarelli et al. (2010, PNAS, 107:2681–2686) used TMS-EEG to measure cortical effective connectivity during midazolam sedation. In waking, a TMS pulse to premotor cortex elicits a complex, spatially differentiated EEG response lasting 300+ ms. Under midazolam, the response is a simple, stereotyped wave that dies out within 150 ms — a loss of cortical complexity. The Perturbational Complexity Index (PCI), developed by Casali et al. (2013, Sci. Transl. Med., 5:198ra105), quantifies this: PCI drops from ~0.44 (waking) to ~0.31 (dreaming) to ~0.18 (propofol NREM-like anesthesia).
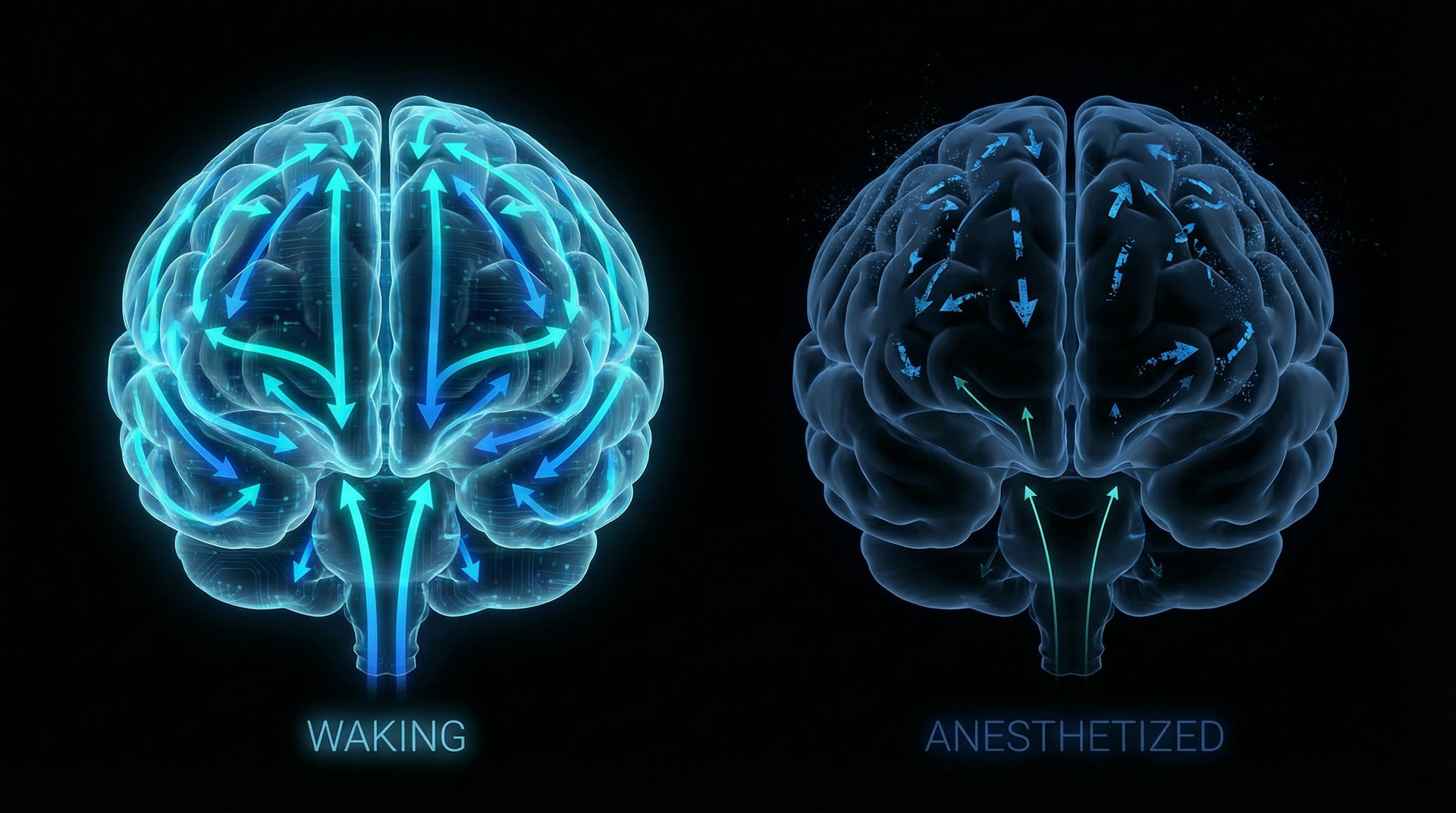
Critically, feedforward connectivity (V1 → V2 → V4 → IT) is preserved under anesthesia while feedback connectivity (IT → V4 → V2 → V1) collapses. Mashour et al. (2020, N. Engl. J. Med., 382:2287–2298) showed that anterior-to-posterior directed information transfer (measured by symbolic transfer entropy, STE) drops by >60% at loss of consciousness, whereas posterior-to-anterior transfer decreases by only ~20%. The brain retains its ability to relay sensory data forward through the hierarchy but loses the top-down predictive signalling that interprets and integrates that data.
Not silence, but disconnection — feedforward preserved, feedback lost
Molecular mechanism: Propofol (2,6-diisopropylphenol) binds to the β₂/β₃ subunits of GABA_A receptors at the transmembrane domain interface (Jurd et al. 2003, FASEB J., 17:250–252). It prolongs the open-time of the chloride channel from ~5 ms (baseline) to ~15–25 ms, approximately tripling the decay time constant of inhibitory postsynaptic currents (IPSCs). At clinical effect-site concentrations of 2–5 µg/mL, this produces progressive cortical and thalamic inhibition.
EEG signature: At the moment of loss of consciousness (LOC), propofol produces a paradoxical increase in frontal alpha power (8–13 Hz) — a 3–10× increase over baseline, clearly visible on raw EEG as large-amplitude (~50–80 µV) rhythmic activity maximal at Fz/Cz electrodes (Purdon et al. 2013, PNAS, 110:E1142–E1151). This "anteriorised alpha" is distinct from waking posterior alpha: it is generated by hypersynchronised thalamocortical loops in which thalamic relay neurons, hyperpolarised by GABA-ergic inhibition from the thalamic reticular nucleus (TRN), switch from tonic firing to T-type Ca²⁺ burst mode at 8–12 Hz.
Slow oscillation: At deeper planes (effect-site concentration >4 µg/mL), propofol induces a slow oscillation (<1 Hz) superimposed on the frontal alpha. Lewis et al. (2012, PNAS, 109:E3377–E3386) showed this reflects cortical UP/DOWN state alternation: during the DOWN state (~400 ms), virtually all cortical neurons are silent — a traveling wave of inhibition sweeps across the cortex at ~1.2–7 m/s. This creates a "fragmentation" of cortical processing — neurons in the DOWN state cannot participate in any computation, even if they are structurally intact.
Connectivity impact: Propofol preferentially disrupts connectivity between the frontal cortex and posterior association areas. Boly et al. (2012, J. Neurosci., 32:7082–7090) showed that propofol anesthesia reduces top-down connectivity (prefrontal → parietal) by 60–70% while leaving bottom-up connectivity relatively intact (~20% reduction). The thalamus is the bottleneck: propofol-induced TRN hyperactivity gates thalamocortical relay, preventing the recurrent frontal-parietal interactions required for conscious processing.
Molecular mechanism: Ketamine is a non-competitive antagonist of NMDA receptors, binding to the phencyclidine (PCP) site within the ion channel pore (Anis et al. 1983, Br. J. Pharmacol., 79:565–575). IC₅₀ for NMDA receptor blockade is ~0.5–1 µM. Clinical plasma concentrations for anesthesia are 9–25 µM (Domino 2010, Anesthesiology, 113:678–684). Ketamine preferentially blocks NMDA receptors on GABAergic interneurons (due to their tonic activation), leading to disinhibition of pyramidal cells — a paradoxical excitation.
EEG signature: Unlike propofol, ketamine does not produce alpha or slow-wave dominance. Instead, it increases broadband gamma power (30–100 Hz) by 200–400% over baseline across frontal and parietal cortex (Akeju et al. 2016, Clin. Neurophysiol., 127:2414–2422). However, gamma coherence — the phase relationship between gamma oscillations in different cortical regions — drops by 40–60%. The brain is oscillating fast but incoherently. Lee et al. (2013, Anesthesiology, 118:1264–1275) showed that ketamine disrupts anterior-to-posterior gamma connectivity more than posterior-to-anterior, the reverse of propofol's effect.
Dissociative phenomenology: At sub-anesthetic doses (0.1–0.5 mg/kg IV), ketamine produces hallucinations, time distortion, and depersonalisation. At full anesthetic doses (1–2 mg/kg IV), patients enter the "K-hole" — a state of complete experiential disconnection. In dynamical systems terms, NMDA-mediated recurrent excitation is essential for maintaining large-scale cortical attractors. When NMDA currents are blocked, the global attractor (a unified conscious state) fragments into multiple local attractors — each cortical region falls into its own semi-independent oscillatory state (Seamans 2008, Pharmacol. Ther., 120:150–162).
A parallel route to this same attractor fragmentation exists through the serotonergic system. Psilocybin — acting primarily as a 5-HT₂A serotonin receptor agonist — produces a strikingly similar dissolution of the unified conscious state. The resulting disruption of thalamocortical integration and default mode network coherence converges on the same network-level endpoint as NMDA blockade: the global attractor fragments, and consciousness enters a dissociated, internally generated state.
Clinical utility: Ketamine preserves airway reflexes and spontaneous respiration at anesthetic doses — distinguishing it from propofol and volatile agents. Heart rate and blood pressure increase by 20–30% due to sympathomimetic effects (centrally-mediated catecholamine release). These properties make ketamine indispensable for anesthesia in hemodynamically unstable patients, field trauma, and resource-limited settings.
Attractor fragmentation — the unified conscious state dissolves into local oscillatory islands
V
Sevoflurane / Isoflurane — The Slow Wave Inducers
Volatile Agent Pharmacology · Burst Suppression · MAC Values · UP/DOWN States
Multi-target pharmacology: Volatile anesthetics (sevoflurane MAC 2.0%, isoflurane MAC 1.15%, desflurane MAC 6.0% — all in O₂ at sea level, age 40) act on multiple molecular targets simultaneously: positive allosteric modulation of GABA_A (prolonging IPSC decay 2–3×), glycine receptor potentiation, NMDA receptor inhibition (IC₅₀ ~0.3–0.5 mM), and activation of two-pore-domain K⁺ channels (TREK-1, TASK) — producing net membrane hyperpolarisation of 5–15 mV in cortical neurons (Hemmings et al. 2005, Trends Pharmacol. Sci., 26:503–510).
Dose-dependent EEG progression: At 0.5 MAC: increased beta activity (paradoxical excitation phase). At 1.0 MAC: dominant slow-delta activity (1–4 Hz, 100–200 µV) with loss of alpha. At 1.5 MAC: burst suppression — alternating periods of high-voltage slow activity (bursts, 1–5 seconds) and isoelectric suppression (flat-line, 2–10 seconds). At >2.0 MAC: prolonged suppression approaching isoelectric EEG (Purdon et al. 2015, Anesthesiology, 123:937–960).
UP/DOWN state physiology: At 1.0 MAC sevoflurane, intracellular recordings in vivo show cortical neurons alternating between depolarised UP states (−55 to −60 mV, sustained firing at 5–20 Hz, duration 200–600 ms) and hyperpolarised DOWN states (−75 to −80 mV, near-complete silence, duration 200–500 ms). The transition from UP to DOWN is mediated by activity-dependent K⁺ channel activation (primarily K_Na/Slack channels); recovery from DOWN to UP depends on I_h and synaptic noise (Compte et al. 2003, J. Neurophysiol., 89:2707–2725). During DOWN states, the cortex is computationally offline — no information processing occurs.
Burst suppression ratio (BSR): At deep anesthesia, the BSR — fraction of time in suppression — is used to titrate anesthetic depth. BSR >50% indicates profound cortical depression. The physiological basis is a global failure of cortical excitability: metabolic demand drops to ~50% of waking levels, and the cortex cycles between brief episodes of self-organised activity (bursts, driven by residual synaptic noise crossing threshold) and periods where excitability is insufficient to sustain any activity (suppression).
Dose-dependent EEG progression — beta to slow-delta to burst suppression
PAC destruction: Theta-gamma PAC is abolished under propofol anesthesia. Mukamel et al. (2014, PNAS, 111:E3283–E3290) measured the modulation index (MI) of theta-gamma coupling in human cortex: MI dropped from 0.018 ± 0.004 (waking) to 0.003 ± 0.002 (propofol LOC) — an 83% reduction. Under sevoflurane at 1.0 MAC, theta is replaced by slow oscillation (<1 Hz), eliminating the scaffold for gamma multiplexing entirely. Under ketamine, gamma persists but its coupling to theta phase becomes random (MI reduced by ~50%, but gamma power increased).
Directed connectivity: Granger causality analysis of multichannel EEG reveals asymmetric connectivity collapse. Ku et al. (2011, PLoS ONE, 6:e25155) showed that propofol reduces feedback (frontal→parietal) Granger causality by 65% while feedforward (parietal→frontal) drops by only 25%. Ketamine shows the opposite asymmetry: Lee et al. (2013) found that ketamine preferentially disrupts anterior→posterior connectivity. Both patterns converge on the same functional consequence: loss of recurrent, integrative processing.
Thalamocortical failure: The thalamus is the convergence point. Under propofol, thalamic relay neurons are hyperpolarised to −70 to −75 mV by enhanced GABAergic input from the TRN, forcing them into burst mode (T-type Ca²⁺ rebound bursts at 8–12 Hz). Under sevoflurane, thalamic neurons are hyperpolarised further (−80 mV) by K⁺ channel activation, entering a tonic silence punctuated by occasional bursts. In both cases, the precise, stimulus-locked thalamic relay required for conveying sensory information to cortex is replaced by rhythmic, uninformative bursting or silence (Alkire et al. 2008, Science, 322:876–880).
The common endpoint: Regardless of molecular mechanism (GABA-ergic, NMDA, multi-target), all general anesthetics converge on a common network-level effect: disruption of the recurrent thalamocortical loops that sustain long-range cortical integration. The diversity of molecular mechanisms producing the same network failure argues that consciousness depends not on any single neurotransmitter system but on the architecture of recurrent connectivity itself.
The thalamus — 1% of brain volume, the critical hub whose disruption abolishes consciousnessThalamocortical recurrent loops — the common target of all anesthetic mechanisms
VII
The Order of Dissolution
Hughlings Jackson Hierarchy · PET/fMRI Data · Association Cortex First
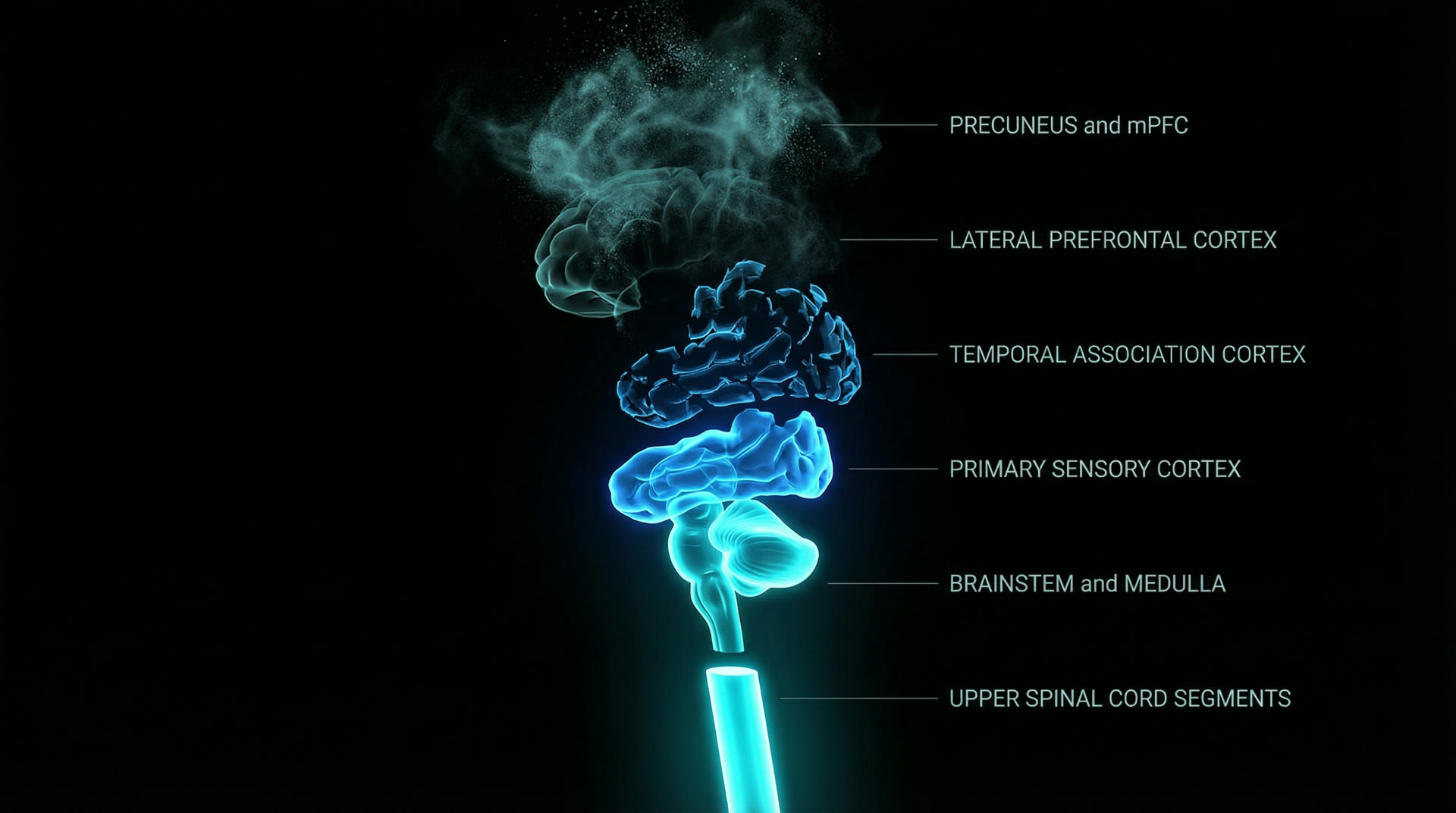
PET studies during propofol induction (Fiset et al. 1999, J. Neurosci., 19:5506–5513) show a characteristic regional sequence of metabolic suppression: first, bilateral precuneus/posterior cingulate cortex (PCC) and medial prefrontal cortex (mPFC) — the default mode network (DMN) hubs. Regional CBF in the precuneus drops by 30–40% before LOC. Second, lateral prefrontal cortex and inferior parietal lobule (the frontoparietal control network). Third, temporal association cortex. Last, primary sensory cortices (V1, S1, A1), which show only 10–15% CBF reduction even at surgical depth.
This hierarchy mirrors Hughlings Jackson's 19th-century principle of dissolution: the phylogenetically and ontogenetically newest structures are the most vulnerable to disruption. The precuneus and mPFC — heavily expanded in the human lineage — are the first to fall. Primary sensory cortex — conserved across all mammals — is the most resistant. Deeper still, the brainstem and medulla — the ancient guardians of respiration, cardiovascular regulation, and arousal — are the last to be suppressed, requiring the highest anesthetic concentrations. The upper spinal cord segments, mediating fundamental motor reflexes and autonomic outflow, remain functional even at surgical planes of anesthesia. Liu et al. (2013, PNAS, 110:13093–13098) confirmed this with functional connectivity analysis: during propofol-induced LOC, precuneus connectivity drops by 50% while V1 connectivity decreases by only 15%.
the exact sequence starts with domino effects like " switch boards shutting off in sequence ONE by ONE " in Hypothalamic Sleep / Wake centres ...... Like Powerhouse tripping Down
The clinical implication is measurable: at sub-anesthetic concentrations (propofol effect-site 1.0–1.5 µg/mL), patients lose self-reflection and autobiographical memory before losing perceptual awareness. At 1.5–2.0 µg/mL, they lose response to verbal command (MOAA/S score 2) while still withdrawing from painful stimuli. At 2.5–4.0 µg/mL, even withdrawal to pain is lost. Each step corresponds to the loss of a progressively more basic cortical function.
This graded dissolution means there exist intermediate "disconnected consciousness" states — where some cortical processing continues (primary sensory, brainstem reflexes) but it is not integrated into unified experience. The brainstem, medulla, and upper spinal cord persist as the irreducible substrate — the structures whose preservation is the anesthesiologist's non-negotiable mandate, for their failure is not unconsciousness but death. Sanders et al. (2012, Anesthesiology, 116:946–959) reported that 37% of patients under general anesthesia have isolated forearm test (IFT) responses suggesting retained (but typically unreportable) cortical processing.
Hughlings Jackson dissolution — newest structures fall first, brainstem · medulla · upper spinal cord persist longest
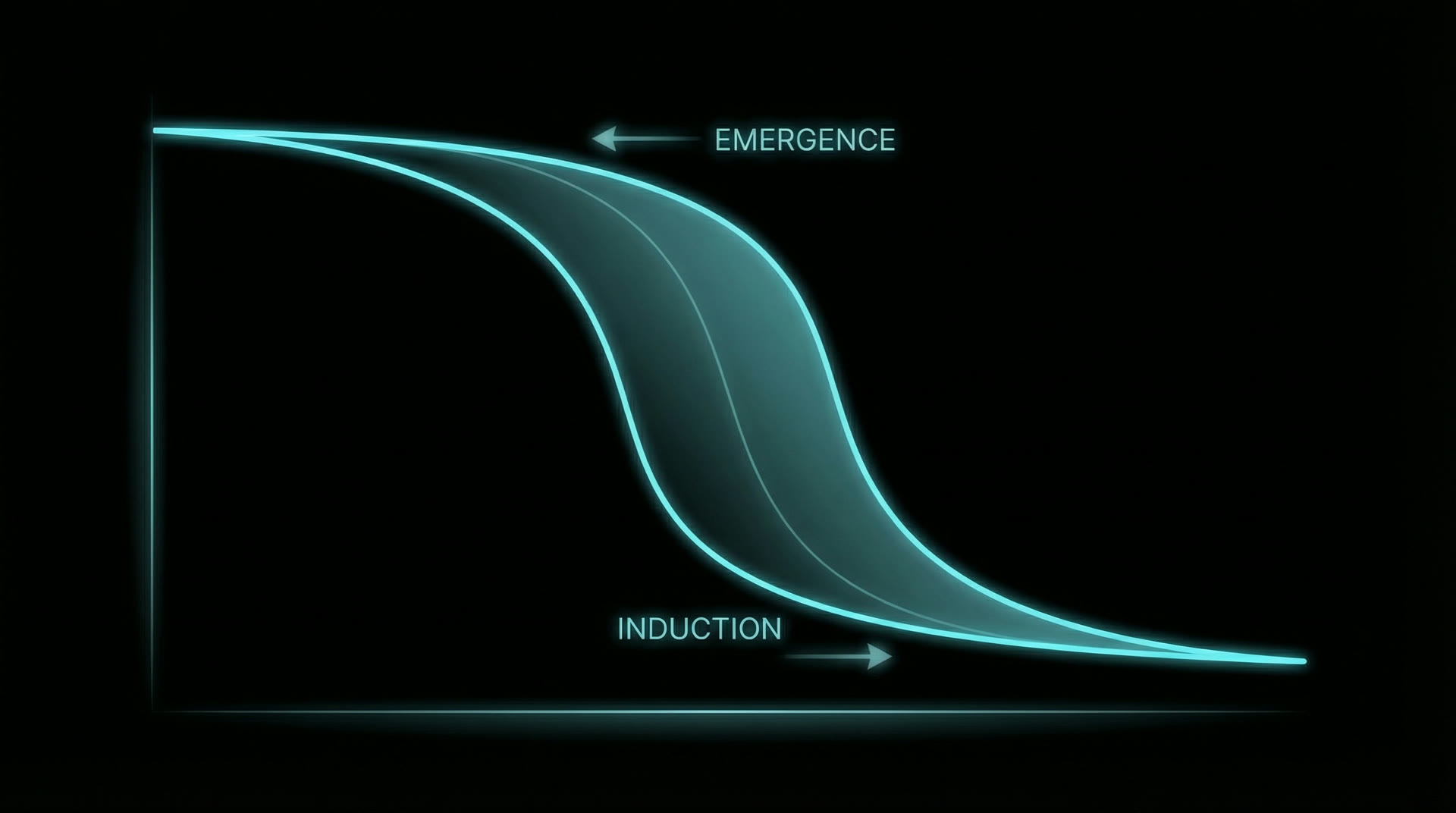
Hysteresis: The anesthetic concentration required for LOC (CE₅₀-LOC for propofol ≈ 3.4 µg/mL) is significantly higher than the concentration at which consciousness returns during emergence (CE₅₀-ROC ≈ 1.6 µg/mL) — a hysteresis ratio of approximately 2:1 (Friedman et al. 2010, Anesthesiology, 112:1056–1066). This is not explained by pharmacokinetic delay alone; it reflects a genuine dynamical hysteresis — the neural state must cross a different bifurcation boundary during recovery than during induction.
Bifurcation model: Steyn-Ross et al. (2004, Phys. Rev. E, 70:011911) modelled LOC as a first-order phase transition in a mean-field cortical model. Below a critical inhibition threshold, the cortex exists in a high-firing "conscious" state. Above threshold, it collapses to a low-firing "unconscious" state. The transition is abrupt (bifurcation), and the return path requires the inhibition to drop below a different (lower) threshold — hence hysteresis. The width of the hysteresis zone depends on the strength of recurrent excitation: stronger recurrence produces wider hysteresis.
Sequential recovery: The recovery order is: (1) brainstem arousal (pupillary reflex, respiratory drive) — mediated by locus coeruleus norepinephrine and dorsal raphe serotonin, which are tonically inhibited under anesthesia and recover first as drug concentration drops; (2) thalamocortical oscillations — alpha and theta rhythms return, visible on EEG as the "alpha return" at propofol CE ~1.5 µg/mL; (3) sensory processing — the patient responds to loud sound or pain; (4) cortical integration — the patient follows commands, indicating frontoparietal network restoration; (5) metacognitive awareness — the patient reports "I'm awake," indicating DMN and prefrontal function (Hight et al. 2014, Br. J. Anaesth., 113:ii49–ii57).
EEG biomarkers of emergence: Purdon et al. (2013) identified specific EEG signatures that predict imminent recovery: (1) return of posterior alpha (8–12 Hz); (2) shift of peak alpha frequency from frontal to occipital; (3) increase in beta power (>15 Hz); (4) restoration of anterior-posterior alpha coherence. These markers can be used for real-time monitoring, enabling anesthesiologists to anticipate emergence 2–5 minutes before behavioral signs.
Hysteresis — consciousness returns at a lower concentration than that which extinguished it
Lesson 1 — Integration, not activity: The brain under propofol at 1.0 MAC generates more rhythmic power (frontal alpha amplitude 50–80 µV) than the waking brain (posterior alpha 30–50 µV). Yet the patient is unconscious. PCI data (Casali et al. 2013) demonstrate that consciousness correlates not with total neural activity but with the complexity of the response to perturbation — the degree to which activity is simultaneously differentiated and integrated. PCI > 0.31 reliably indicates consciousness; PCI < 0.31 indicates unconsciousness, across all tested anesthetic agents, sleep stages, and vegetative/minimally conscious patients.
Lesson 2 — Feedback connectivity is necessary: Across all agents studied (propofol, sevoflurane, ketamine, xenon), the most consistent neural correlate of unconsciousness is the loss of top-down (feedback) directed connectivity, measured by Granger causality, directed transfer function, or symbolic transfer entropy (Mashour 2014, Curr. Opin. Anaesthesiol., 27:478–482). Feedforward processing is preserved. This supports predictive coding models: consciousness requires the brain to generate and test predictions (feedback), not merely to relay sensory data (feedforward).
Lesson 3 — THE THALAMUS IS THE CRITICAL HUB: Every major anesthetic mechanism converges on the thalamus. Propofol: TRN-mediated hyperpolarisation of relay neurons. Ketamine: disruption of corticothalamic glutamatergic feedback. Sevoflurane: K⁺-channel-mediated tonic hyperpolarisation. Dexmedetomidine (α₂-agonist): direct inhibition of locus coeruleus → thalamic deafferentation. The thalamus, comprising only ~1% of total brain volume, is the necessary relay station whose disruption most reliably abolishes consciousness (Alkire et al. 2008).
Lesson 4 — Consciousness is graded, not binary: Sedation scales (MOAA/S, BIS, Patient State Index) reveal a continuum. BIS values: 90–100 (awake), 60–80 (sedated, may respond to loud voice), 40–60 (general anesthesia, no response to command, suppressed cortical integration), <40 (deep anesthesia, burst suppression). There is no single "consciousness switch" — there is a progressive loss of increasingly basic neural functions as anesthetic depth increases.
Perturbational Complexity — waking brain responds with rich differentiation, anesthetised brain with stereotyped simplicity
X
Vedantic Parallel
Māṇḍūkya Upaniṣad · Four States · A Brief Philosophical Note
The Māṇḍūkya Upaniṣad describes four states of consciousness — Vaiśvānara (waking, externally directed), Taijasa (dreaming, internally generated), Prājña (deep dreamless sleep, undifferentiated awareness), and Turīya (the fourth, substratum of all three).
The first two states map well to anesthesia observations:
However, Prājña and Turīya require caution in this mapping. These are states achieved through deliberate, voluntary meditative effort — not pharmacologically induced unconsciousness. The deep dreamless sleep of Prājña and the transcendent substratum of Turīya belong to a different phenomenological domain than anesthetic suppression. The parallel is suggestive but not yet sufficiently developed to be presented as equivalence. This remains an area for further exploration.
This mapping is descriptive, not explanatory. The scientific value of anesthesia research lies in its quantitative, mechanistic findings — specific receptor affinities, measurable EEG signatures, directed connectivity metrics, and dose-response relationships. The Upanishadic framework provides a phenomenological taxonomy that has endured for ~2,800 years, suggesting it captures something real about the structure of conscious states — but the explanatory work is done by the neuroscience.
As a cardiac anesthesiologist with over four decades of clinical practice, the author has observed these state transitions thousands of times: the precise moment a patient's eyes close under propofol induction (typically at effect-site concentration 2.5–3.5 µg/mL), the characteristic EEG shift from posterior alpha to frontal alpha to slow-delta, the graded recovery sequence during emergence. These observations are not philosophical abstractions. They are repeatable, measurable, and increasingly predictable — the hallmarks of genuine science.